



Category Archives: evenements
Séminaire IGN/AgroParisTech/LabEx ARBRE
Séminaire IGN/AgroParisTech/LabEx ARBRE
Ressources forestières et halieutiques : dynamique, modélisation, et appui aux politiques publiques dans un contexte incertain et contraint
14 octobre 2021 à 9h30
Salle de conférence
Centre INRAE-Champenoux
Ce séminaire scientifique est organisé par l’IGN et AgroParisTech dans le cadre de la soutenance de thèse de Timothée Audinot : Développement d’un modèle de dynamique forestière à grande échelle pour simuler les forêts françaises dans un contexte non-stationnaire. Thèse de doctorat Université de Lorraine, école doctorale SiReNa, Direction : J-D Bontemps (IGN), H Wernsdörfer (AgroParisTech).
Inscription et passe sanitaire requis

EST’ival des Sciences
L’association Communicasciences a organisé pour la première fois cette année l’EST’ival des Sciences : une tournée de 20 événements de culture scientifique dans tout le Grand Est, jusque dans les petites communes, du 09 au 31 juillet 2021 à raison de 2 interventions par département. Les interventions ont été réalisées par des scientifiques issus de laboratoires de recherche publique de la région, accompagnés par des médiateurs de l’association afin d’aborder une thématique de recherche par commune. Les scientifiques du LabEx ARBRE ont participé à cet événement : Francis Martin est intervenu à Doulaincourt-Saucourt en Haute-Marne et Irène Carravieri & Sandrine Capizzi sont intervenues dans le cadre de l’initiative CiTIQUE à Verzenay dans la Marne.


CiTIQUE : convention partenariale entre INRAE et le CNPF
Ce vendredi 3 septembre, nous accueillerons sur le Centre à Champenoux entre 12h et 16h, Philippe Mauguin (Président-Directeur Général d’INRAE) et Claire Hubert (Directrice Générale du CNPF) pour la signature de la convention partenariale entre INRAE et le CNPF autour du programme CiTIQUE, en la présence de Nicole Trisse (Députée de la 5ème circonscription de Moselle), Dominique Potier (Député de la 5ème circonscription de Meurthe et Moselle) et Pierre Mutzenhardt (Président de l’Université de Lorraine).
Ce partenariat doit permettre de constituer une source importante de données, utiles aux chercheurs d’INRAE et aux autres laboratoires de recherche partenaires de CiTIQUE, pour approfondir leur connaissance sur l’écologie des tiques et pour élaborer des modèles de risque. Cette collaboration est nouée aussi dans le but de consolider la politique de prévention mise en place par le CNPF vis-à-vis de ses agents exposés au risque posé par les maladies vectorielles à tiques puisqu’ils pourront signaler les piqûres de tiques survenues dans le cadre de leurs activités professionnelles.
Soutenance HDR
Laure VIEUBLE GONOD soutiendra son HDR intitulée : “Effets des propriétés indigènes du sol et des activités anthropiques sur les microorganismes du sol et leurs activités en contexte agricole” le vendredi 12 mars à partir de 9h devant le jury composé de : Dominique PATUREAU (rapportrice), Sylvie RECOUS (rapportrice), Lionel ALLETTO (rapporteur), Claire CHENU (examinatrice) et Fabrice MARTIN LAURENT (examinateur)
Elle se déroulera complètement en distanciel et sera accessible avec le lien suivant :
https://eu.bbcollab.com/guest/cac0251ff3254c46a14f4c759bb65074
Vous trouverez ci-dessous un résumé des travaux qui seront présentés :
Les organismes du sol et en particulier, les microorganismes du sol, jouent un rôle clef dans le fonctionnement des sols et sont impliqués dans de nombreux services écosystémiques. La biodiversité des sols a besoin d’être davantage onnue afin que l’on puisse mieux la diagnostiquer et la piloter dans le but de restaurer, maintenir et améliorer les services rendus par les sols. Les recherches développées jusqu’à aujourd’hui ont donc consisté à mieux comprendre le fonctionnement microbiologique des sols (et notamment la biodégradation) dans un contexte agricole. Le fonctionnement microbiologique des sols est contrôlé par les propriétés indigènes du sol mais peut également être impacté par les activités anthropiques et notamment les pratiques agricoles. Plus spécifiquement, mes activités de recherches, qui se déclinent à différentes échelles spatiales (du µm à la parcelle) ont porté sur l’impact de i) la structure du sol sur les microorganismes du sol et leurs activités et ii) des pratiques agricoles sur les microorganismes et leurs activités (biodégradation et devenir des pesticides dans les sols, recyclage des produits résiduaires en agriculture).
Pour ce faire, j’ai mis en oeuvre des approches essentiellement expérimentales mais j’ai également participé occasionnellement à des travaux de modélisation. Mes travaux sont à la fois fondamentaux et mécanistes mais également finalisés, ces derniers ayant pour objectifs d’apporter des éléments de réponses vis-à-vis de questions sociétales (polluants émergents, valorisation des déchets…). Mon projet de recherche pour les années à venir vise à approfondir les thématiques de recherche initiées tout en diversifiant les contextes (rural, périurbain, urbain) et en considérant d’autres systèmes de culture (alternatifs vs conventionnels) en lien avec des demandes sociétales et politiques fortes (besoin de nature en ville, développement de l’agroécologie, changement climatique…) et à prendre en considération les autres organismes du sol et en particulier la faune pour développer des connaissances sur les réseaux trophiques et les relations diversité-fonctions-services écosystémiques.
Mots clés : sol, bactéries, champignons, distribution spatiale, minéralisation, accessibilité, diffusion, advection, pesticides, produits résiduaires organiques
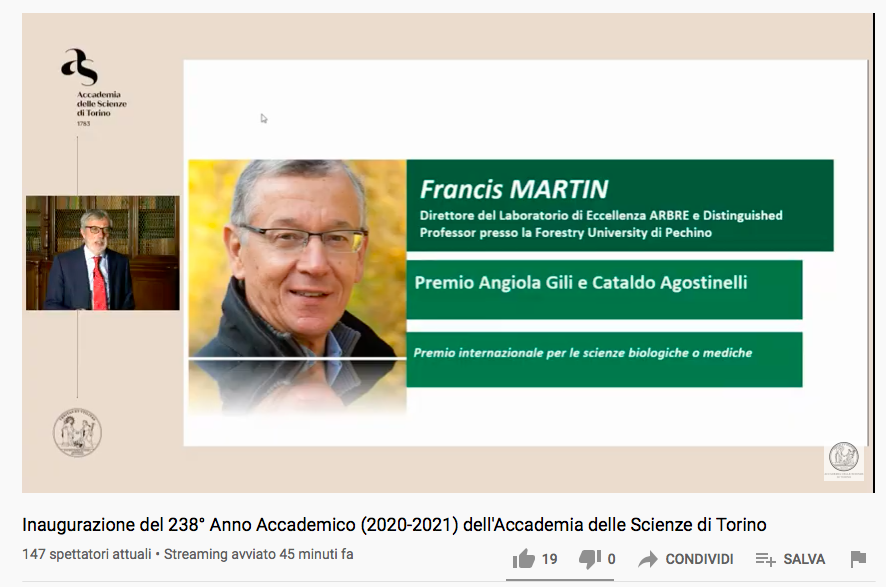
Prix Gili Agostinelli 2020 : une reconnaissance internationale décernée à Francis Martin
Le 1er mars dernier, Francis Martin, directeur de recherche INRAE au sein du laboratoire IAM (Interactions Arbres/Micro-organismes) et directeur du LabEx ARBRE s’est vu décerner le prix international Gili Agostinelli 2020 par l’Académie des Sciences de Turin. Une reconnaissance internationale de ses travaux de recherche sur la biologie et l’écologie des champignons forestiers.
La remise du Prix Gili Agostinelli s’est déroulée lors de la Cérémonie d’ouverture de l’année académique 2020-21 de l’Académie des sciences de Turin le 1er mars dernier. Les conditions sanitaires n’ayant pas permis un événement sur place, la cérémonie a été diffusée en ligne. Le Président de l’Académie a précisé dans son discours de remise du prix : “Francis Martin a développé des recherches innovantes sur la biologie et l’écologie des champignons forestiers, basées sur la génomique comparative à grande échelle. Ces travaux ont conduit à plusieurs publications dans les revues scientifiques prestigieuses (Nature, Science, PNAS). Le séquençage du génome du champignon symbiotique Laccaria bicolor, suivi de celui de la Truffe noire du Périgord, tous deux publiés dans la revue scientifique Nature (2008, 2010), ont constitué une avancée majeure dans la compréhension du mode de vie des champignons symbiotiques. Avec ses contributions (de la génomique comparative à la description des molécules de signalisation fongique), Francis Martin a fait émerger les symbioses mycorhiziennes du domaine des spécialistes dans lequel elles étaient reléguées depuis les années 70. Il nous a fait comprendre leur importance tant pour la biologie fondamentale, comprise comme la connaissance des processus de la vie, que pour la bonne santé des écosystèmes. Au-delà des avancées qu’il a réalisées en génomique (du peuplier, le premier arbre séquencé, aux champignons symbiotiques et pathogènes), il faut rappeler d’autres travaux majeurs, comme l’identification chez les champignons mycorhiziens, de protéines de communication contrôlant l’interaction entre les champignons symbiotiques et leurs arbres-hôtes.
Grâce à sa notoriété, son parcours scientifique et les résultats de ses recherches, Francis Martin représente un modèle d’excellence scientifique, capable de décrypter avec les outils de la génomique les processus écologiques et biologiques qui sont à la base du bon fonctionnement de notre planète.”
—–
Francis Martin fait partie des cinq chercheurs lorrains « hautement cités » en 2020, distinction prestigieuse suite à la publication par la société Clarivate Analytics d’une liste des chercheurs ayant fait preuve d’une influence internationale notable à travers le nombre de publications hautement citées.
Rapport sur l’efficacité et le financement de la lutte contre la maladie de Lyme
Le Labex ARBRE est porteur avec INRAE du programme national de recherche participative CiTIQUE (pour « citoyens » et « tiques ») qui s’attache à observer, surveiller et prévenir les risques liés aux tiques. Ce programme fait appel aux citoyens dans toute leur diversité (grand public, élèves, enseignants, jardiniers, randonneurs, promeneurs, forestiers, agriculteurs, vétérinaires, médecins, élus,…), pour participer à l’effort de recherche dans la lutte contre les maladies vectorielles à tiques. Il permet aussi aux citoyens de s’impliquer aux côtés des chercheurs à toutes les étapes de la démarche scientifique lors de stages au laboratoire Tous Chercheurs de Nancy (https://www.touschercheurs.fr/sciences-participatives/nancy/citique/), dans un climat d’écoute mutuelle et de partage des savoirs.
Le 3 mars dernier, Mme la Député Louwagie a présenté en séance publique à l’Assemblée Nationale les conclusions de son rapport sur l’efficacité et le financement de la lutte contre la maladie de Lyme, dans lequel elle souligne la réussite de CiTIQUE et recommande de « consolider et étendre le projet CiTIQUE visant à surveiller, de manière participative, l’exposition de la population aux tiques ». C’est une belle reconnaissance des efforts engagés par INRAE, le Labex et le Centre Permanent d’initiatives pour l’Environnement de Nancy Champenoux pour faire vivre le programme CiTIQUE, et cela souligne aussi toute l’importance de la recherche participative en matière de santé publique !

Carbone dans les sols : mise au point d’un modèle mathématique pour mieux comprendre le cycle de la matière organique
Le sol, là où tout commence et se termine. La matière organique du sol est une véritable clé de voûte de la sécurité alimentaire, de la biodiversité ou encore de l’atténuation du changement climatique. Les progrès techniques récents en sciences du sol et en microbiologie ont permis de mettre en évidence des processus de décomposition de la matière organique jusqu’alors insoupçonnés. Des chercheurs d’INRAE, de l’Université de Lorraine et d’AgroParisTech se sont associés pour développer un modèle mathématique innovant, qui intègre ces découvertes récentes. Leurs travaux financés par le LabEx ARBRE, sont publiés Nature Communications du 5 février 2021. Ils apportent un éclairage nouveau sur le cycle de la matière organique du sol.
Le sol, et plus particulièrement la matière organique (MO) qui le compose est de plus en plus au cœur des travaux de recherche, en raison de sa capacité à piéger le carbone atmosphérique et de son rôle majeur dans la santé des sols. La MO est composée de molécules carbonées très variées. De multiples acteurs microscopiques, les micro-organismes, agissent de concert pour la décomposer à l’aide de leurs outils que sont les enzymes. La compréhension de la dynamique de décomposition de la MO a considérablement progressé au cours des dernières années. Jusqu’à présent la MO était considérée comme un ensemble de composés difficiles à dégrader. Aujourd’hui, on considère que cette MO est faite d’un ensemble de molécules qui sont continuellement transformées par les micro-organismes pour devenir de plus en plus petites. Cette nouvelle vision remet en question les modèles existants de prédiction de la dynamique du carbone.
C’est pourquoi des chercheurs d’INRAE, de l’Université de Lorraine et d’AgroParisTech se sont associés, pour développer un modèle qui prend en compte les nouvelles données en chimie du sol et écologie microbienne. Baptisé C-STABILITY, ce modèle combine les approches mathématiques des différents modèles actuels et reproduit avec succès les processus de la dynamique de la MO. C-STABILITY traduit les transformations réalisées par les enzymes et les microbes du sol tout en mettant l’accent sur l’accessibilité de la MO.
Les simulations théoriques réalisées grâce à ce nouveau modèle mathématique apportent un éclairage nouveau sur la relation entre les micro-organismes décomposeurs, la chimie de la MO et le stock de MO. La structure mathématique flexible de C-STABILITY offre une base prometteuse pour l’exploration de nouvelles hypothèses de recherche sur la MO et pour améliorer l’évaluation de la capacité des sols à stocker le carbone.
Référence
Sainte-Marie, J., Barrandon, M., Saint-André, L. et al. C-STABILITY an innovative modeling framework to leverage the continuous representation of organic matter. Nat Commun 12, 810 (2021). https://doi.org/10.1038/s41467-021-21079-6
Photo : N. Pousse (ONF)

|
|
Production contrôlée de truffes blanches made in France
Tuber magnatum Pico, connue sous le nom de Truffe blanche du Piémont ou Truffe blanche d’Italie, est la truffe la plus rare et la plus chère. Elle est récoltée exclusivement en forêt dans quelques pays d’Europe, mais l’offre n’arrive souvent pas à combler la forte demande mondiale pour ce champignon. Depuis 2008, après 9 ans de recherches conjointes entre INRAE et les pépinières ROBIN, les premières plantations truffières ayant pour but sa culture ont pu être faites en France. La persistance de la truffe blanche a été vérifiée dans des truffières ayant de trois à huit ans, et dans l’une d’elles, âgée de quatre ans, les premières récoltes ont eu lieu en 2019. Les résultats scientifiques de ces travaux sont parus le 16 février dans la revue Mycorrhiza. La production de fructifications de T. magnatum dans une plantation hors de son aire de distribution naturelle est une première mondiale ouvrant la voie au développement de la culture de cette truffe en France mais aussi ailleurs dans le monde.
La truffe blanche italienne (Tuber magnatum Pico) est la star des truffes pour de nombreux grands restaurants gastronomiques à travers le monde. Son parfum particulier la rendait déjà unique dans les années 1700, alors que les princes de Savoie l’utilisaient dans leurs négociations diplomatiques. La fructification (truffe) de T. magnatum est produite par un champignon qui vit en symbiose avec des arbres tels que les chênes, les saules, les charmes et les peupliers. Cette truffe est récoltée naturellement en Italie, dans la péninsule balkanique, plus rarement en Suisse et dans le sud-est de la France. La production annuelle de cette truffe est de quelques dizaines de tonnes.
Depuis 2008, la pépinière ROBIN commercialise des arbres mycorhizés par T. magnatum suivant le procédé INRAE/pépinières ROBIN, sous licence et contrôle d’INRAE. Chaque plante est ainsi vérifiée individuellement avant sa commercialisation par des experts INRAE, qui contrôlent la présence de la truffe en analysant des caractéristiques morphologiques et effectuant des analyses ADN. Dans un programme de recherche conjoint INRAE/pépinières ROBIN, cinq plantations françaises ont été étudiées. Le premier résultat est la persistance dans le sol trois à huit ans après plantation de la truffe blanche pour quatre plantations réparties dans des régions au climat différent (Rhône-Alpes, Bourgogne Franche Comté et Nouvelle Aquitaine). Le principal résultat de ce travail a été la récolte en 2019 de trois truffes et quatre en 2020 dans la plantation de Nouvelle-Aquitaine. Ces truffes sont ainsi les premières récoltées dans une plantation en dehors de l’aire de répartition géographique naturelle de cette espèce.
La trufficulture connaît un essor mondial depuis quelques années. En France, elle se développe également dans de nombreuses régions en permettant aux agriculteurs de se diversifier tout en respectant l’environnement : c’est une culture agroécologique ne nécessitant pas d’intrants chimiques et favorisant la biodiversité. Les résultats de cette étude ouvrent la voie à la culture de T. magnatum hors de sa zone de distribution naturelle, à condition de planter des plants mycorhizés de haute qualité dans des sols adaptés et d’appliquer une gestion appropriée des plantations.
Cette première mondiale a été saluée par Joël Giraud, secrétaire d’État auprès de la ministre de la Cohésion des territoires et des Relations avec les collectivités territoriales, chargé de la Ruralité.
Référence
Bach, C., Beacco, P., Cammaletti, P. et al. First production of Italian white truffle (Tuber magnatum Pico) ascocarps in an orchard outside its natural range distribution in France. Mycorrhiza (2021). https://doi.org/10.1007/s00572-020-01013-2

Ateliers REGEFOR 2021
Du fait de l’incertitude des conditions sanitaires d’ici le mois de juin, le comité organisateur a pris la décision de basculer les ateliers ReGeFor 2020 (21-23 juin 2021) en format 100% virtuel. Le site internet dédié à l’événement est mis régulièrement à jour. (https://journees.inrae.fr/regefor/ ).
